La storia della genomica: come nasce, come si sviluppa e cosa ci riserva in futuro
di Sergio Barocci – Università di Genova per la terza età e divulgatore scientifico.
Dalla nucleina alla strutturistica di Watson e Crick (parte I)

La genomica moderna come oggi la conosciamo, in realtà ha avuto solo inizio nel 1970. anche se le sue fondamenta siano state in precedenza costruite con diverse pietre miliari significative. Infatti, nell’arco di circa 150 anni si è assistito a una vera e propria rivoluzione in campo biologico, che ha permesso di far luce su uno degli aspetti più affascinanti e misteriosi della vita cioè il patrimonio genetico degli organismi, dalla sua composizione alla sua struttura sino ad arrivare al suo funzionamento.
Ripercorriamo queste tappe partendo partendo dal biologo svizzero Friedrich Miescher (1844–1895) che pubblicò nel 1871 un articolo in cui sosteneva di aver identificato la presenza di nucleina e proteine associate nel nucleo delle cellule. La scoperta della nucleina, infatti, fu il primo passo per l’identificazione del DNA come molecola responsabile della trasmissione dei caratteri genetici e la base del campo della genomica.
Albrecht Kossel e la teoria cromosimica

Tra il 1885 e il 1901 il chimico tedesco Albrecht Kossel (1853–1927) incentrò i suoi studi sui processi biochimici delle proteine, ed in particolare sull’urea e sui derivati degli acidi nucleici giungendo ad isolare e descrivere cinque composti organici presenti nell’acido nucleico: adenina (A), citosina (C), guanina (G), timina (T), uracile (U), oggi noti per fornire la struttura molecolare nella formazione del DNA e dell’ RNA.
Nel 1904, il biologo statunitense Walter Stanborough Sutton (Utica, 5aprile 1877 – Kansas City, 10novembre 1916) ed il biologo tedesco Theodor Heinrich Boveri (1862 – 1915) motivarono la teoria cromosomica dell’ereditarietà: i cromosomi rappresentano gli “elementi organizzati autonomamente nella cellula” e passano da questa alle cellule figlie e da un individuo alla sua discendenza.
Nel 1910 il biochimico lituano Phoebus Levene (1869–1940), emigrato nel 1893 negli USA a causa di una sommossa antisemita che aveva come bersaglio anche la sua famiglia, di origine ebraica, fu il primo a scoprire l’ordine delle tre componenti principali di un singolo nucleotide (fosfato – zucchero – base azotata) ma anche il primo a scoprire nel 1909 il componente in carboidrati dell’RNA (il d-ribosio) e nel 1929 il componente in carboidrati del DNA (il deossiribosio). Propose la struttura del tetranucleotide in cui nucleotidi erano disposti secondo un preciso ordine ripetuto cioè G-C-T-A-G-C-T-A e così via.
da LEVENE a Schrödinger

Tale ipotesi, prima della formulazione di Erwin Chargaff, ebbe un successo tale da portare la maggior parte della comunità scientifica a non credere che il DNA potesse essere vettore dell’informazione genetica.
In ogni caso, il lavoro di Levene divenne fondamentale per delineare la vera struttura del DNA, struttura che egli non poté tuttavia apprezzare, a causa della sua morte avvenuta nel 1940, prima che il suo vero significato diventasse chiaro.
Nel 1937 il fisico inglese William Astbury (Longton, 25 febbraio 1898 – Leeds, 4 giugno 1961) presentò i primi risultati di alcuni studi di diffrazione ai raggi X che dimostrarono una struttura estremamente regolare del DNA e nel 1944 si dovette al fisico e matematico Erwin Schrödinger (Vienna, 12agosto 1887 – Vienna, 4gennaio 1961), vincitore del Premio Nobel per la Fisica nel 1933, l’ipotesi del “cristallo aperiodico” cioè che il materiale genetico, secondo la fisica quantistica, doveva essere costituito da una molecola di grandi dimensioni dalla struttura non ripetitiva, sufficientemente stabile da mantenere l’informazione genetica.
Griffith ed il principio trasformante

Qualche anno prima, attraverso alcuni esperimenti si gettarono le basi che il DNA costituiva la sede dell’eredità biologica.
Nel 1928 fu il biologo inglese Frederick Griffith (1879 -1941) a dimostrare che ceppi di Diplococcus pneumoniae (il cui nome scientifico attuale è quello di Streptococco pneumoniae) uccisi al calore potevano trasformare un ceppo non virulento in virulento. Tale esperimento aprì la strada alla determinazione della natura del materiale genetico. Tale scoperta rappresentò uno dei primi esperimenti che suggerirono come i batteri fossero in grado di trasferire le informazioni genetiche attraverso un processo noto come trasformazione.
Questo sistema, pur non fornendo nessuna evidenza su quale fosse la sostanza che determinava il cambiamento, mostrò che qualcosa poteva trasportare l’informazione genetica dai resti dei batteri morti a quelli vivi.
l’esperimento del dott. Avery

Si parlò quindi di un principio trasformante in grado di modificare i batteri vivi.
Nel 1943 il medico canadese Oswald T. Avery (1877–1955) ma anche uno dei primi biologi molecolari e pioniere nel campo dell’immunologia, dimostrò in un celebre esperimento insieme a suoi collaboratori Colin MacLeod (1909–1972) e Maclyn McCarty (1911–2005) che il DNA era il principio trasformante alla base di questo fenomeno.
Tale esperimento, che si basava su quello di Griffith, diede una chiara prova del fatto che i geni e i cromosomi erano costituiti da DNA. Griffith e colleghi riuscirono a lisare i pneumococchi e separarne le componenti macromolecolari (proteine, polisaccaridi, lipidi ed acidi nucleici). Le cavie sopravvivevano quando trattate con tutte le biomolecole tranne gli acidi nucleici: il materiale genetico trasformante doveva essere quindi DNA e/o RNA.
ignorati fino alla morte

La comunità scientifica fu riluttante ad accettare i risultati di Avery. Infatti, per molti anni fino al 1952, il suo lavoro fu ignorato e Avery morì nel 1955 senza aver ricevuto il premio Nobel che probabilmente meritava.
Nel frattempo, grazie alle scoperte di Levene, in considerazione della presenza nell’RNA di un pentoso (ribosio) con un OH in posizione 2’ (che impedisce a sequenze complementari di assumere una struttura a doppia elica del tipo B del DNA, conferendo una maggiore disponibilità a interazioni terziarie e a reazioni chimiche anche del tipo enzimatiche), come anche di una base azotata l’uracile, diversa dalla corrispondente pirimidina (la timina del DNA) si arrivò a distinguere il DNA dall’RNA. Si dovette però aspettare il 1950, con il lavoro del biochimico scozzese John Masson Gulland (14ottobre 1898 – 26ottobre 1947), per proporre in maniera esplicita l’esistenza dentro le cellule viventi di due diversi acidi nucleici.
1950 – 1968: Sviluppo delle conoscenze sul DNA

Così come per Avery, il merito delle scoperte di Gulland verrà riconosciuto solo in maniera postuma.
Tra il 1950 e il 1968 incominciarono a svilupparsi le conoscenze sul DNA. Infatti, fu agli inizi degli anni ‘50 che il biochimico austriaco Erwin Chargaff (1905 -2002) cercò di comprendere meglio le peculiarità del DNA.
Quali erano le caratteristiche chimiche che permettevano a questa molecola di contenere le informazioni genetiche? Questo divenne uno dei primi importanti passi verso la comprensione della struttura dell’acido desossiribonucleico. Chargaff sviluppò la comprensione dell’accoppiamento delle basi nucleotidiche di adenosina, citosina, guanina e timina che diedero un importante contributo nella determinazione della struttura della molecola di DNA evidenziata poi nel 1953 da J. Watson e F. Crick.
Chargaff scoprì, infatti, che c’erano sempre quantità uguali di timina e adenina o citosina e guanina nei campioni di DNA. Di conseguenza, dedusse che l’adenosina formava coppie cromosomiche con la timina e la citosina coppie cromosomiche con la guanina.
DNA COME PRINCIPIO TRASFORMANTE

Si arrivò così alla vigilia della scoperta della doppia elica del DNA: nel febbraio del 1944 O. Avery e collaboratori. avevano pubblicato sulla Rivista Journal Experimental Medicine il loro classico lavoro in cui si dimostrava che il principio trasformante era il DNA. Insieme a scoperte successive, il loro lavoro costituiva una prova sperimentale che il DNA e non le proteine rappresentava la molecola biologica dell’ereditarietà segnando l’inizio dell’era della biologia molecolare. Anche se tale lavoro ebbe poco credito nella comunità scientifica di allora, l’importanza del DNA nel meccanismo dell’ereditarietà divenne totale attraverso un altro classico esperimento detto del “frullatore” da parte dei genetisti statunitensi Alfred Day Hershey (Owosso, 4dicembre 1908 – Syosset, 22maggio 1997) e di Martha Cowles Chase (Cleveland Heights, 30novembre 1927 – Lorain, 8agosto 2003) attraverso lo studio di batteri e batteriofagi.
Esperimento di Hershey-Chase

Tale esperimento, a detta dello stesso A. Hershey, non rappresentava di certo l’esperimento migliore ma uno tra i tanti che consacrarono il DNA come la molecola portatrice dell’ereditarietà biologica. Anche se era stato stabilito che il DNA era il materiale genetico di tutti gli organismi, ad eccezione di alcuni virus, ed erano stati forniti alcuni dettagli su come alcuni componenti chimici di base costituivano gli acidi nucleici, rimaneva ancora da decifrare la struttura precisa del DNA, cioè come venivano organizzate le catene polinucleotidiche in DNA che funzionava da materiale genetico.
Il DNA era composto da una singola catena o da più di una? Se il DNA era formato da due catene come si correlavano chimicamente una catena all’altra, come si diramavano le catene e come correlava la struttura del DNA alle varie funzioni come espressione, replicazione e mutazione?
ALLA RICERCA DELLA STRUTTURA DEL DNA

A questo punto rimaneva ancora aperta un’ultima questione: “Di cosa era fatto il materiale genetico degli “organismi” che non contenevano DNA (i virus a RNA)?”. La risposta a questa domanda non tardò ad arrivare in seguito agli studi condotti sul virus del mosaico del tabacco (TMV) da Alfred Gierer (15aprile 1929 – ) e G. Schramm nel 1956 e da Heinz Fraenkel-Conrat (1910 -1999) e B. B. Singer nel 1957.
Tra il 1949 e il 1953 oltre a E. Chargaff altri scienziati si interessarono a risolvere la struttura del DNA. Tra questi anche la cristallografa britannica Rosalind Franklin (1920–1958), il chimico statunitense Linus Pauling (1901–1994), il biologo neozelandese Maurice Wilkins (1916 – 2004), il biologo statunitense James Watson (1928 – ) e il fisico britannico Francis Crick (1916–2004) che cercarono una qualche informazione che potesse rispondere a quella che molti consideravano la questione più significativa: come faceva il DNA a funzionare da base genetica per i processi vitali? Si ritenne che la risposta dipendesse fortemente dalla struttura chimica e dall’organizzazione della molecola di DNA.
LA DOPPIA ELICA

Gli sforzi furono premiati quando nel 1953 Watson e Crick avanzarono la loro ipotesi sulla natura a doppia elica del DNA, senza fare alcun esperimento ma costruendo dei modelli (in cartone e filo di ferro). Fu provata la correttezza dell’assunzione che le funzioni del DNA sarebbero state più semplici da chiarire una volta determinata la sua struttura generale. Infatti, fino al 1944 alcune osservazioni favorivano le proteine come materiale genetico.
Watson e Crick presentarono sulla rivista Nature del 25aprile del 1953, quello che oggi viene accertato come il primo modello accurato della struttura del DNA, ossia il modello della doppia elica e a disegnarne il bozzetto fu la moglie di F. Crick, la pittrice Odile Speed (King’s Lynn, 11agosto 1920 – San Diego, 5luglio 2007).
I dati sperimentali cruciali per lo sviluppo del loro modello provenivano soprattutto da due fonti: i rapporti quantitativi tra le basi azotate e gli studi di diffrazione a raggi X del DNA.
IL MODELLO DEL DNA

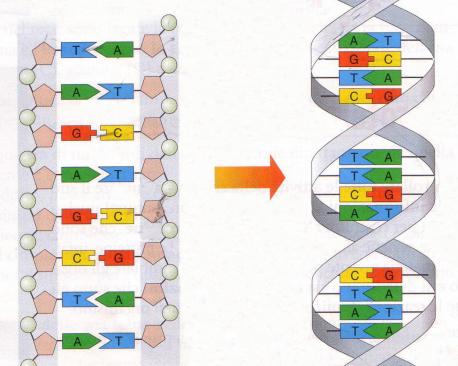
Infatti, i due scienziati ipotizzarono che la molecola del DNA fosse costituita da una doppia elica, cioè da due catene polinucleotidiche spiralizzate con avvolgimento destrorso attorno a un unico asse, connesse tra di loro da legami-idrogeno. Essi proposero che i legami-idrogeno connettessero tra loro le coppie di basi note per essere presenti in quantità uguali, cioè A con T e C con G.
Le risultanti strutture risultano planari, perpendicolari all’asse e di dimensioni identiche (garantendo quindi un diametro costante per la doppia elica, indipendente dalla sequenza delle basi). Le coppie di basi sono complanari; i desossiribosi giacciono su piani perpendicolari a quelli delle basi. Le catene pentoso – fosfato – pentoso – fosfato… sono fortemente anioniche e si devono trovare all’esterno della molecola a contatto con l’acqua, mentre le coppie di basi, idrofobiche, si impaccano le une sulle altre al centro della molecola, escludendo l’acqua.
la struttura del dna proposta da Watson e Crick

Avvicinando due filamenti di DNA c’è repulsione causata dalle cariche negative. Tra le basi che si fronteggiano si formano dei legami idrogeno:
1. Due tra A e T;
2. Tre tra C e G.
Se prevalgono le citosine e le guanine, il numero di legami idrogeno è molto maggiore che se prevalgono adenine e timine. Quando prevalgono C e G il DNA si denatura ad una temperatura maggiore. Tra le basi si instaurano interazioni idrofobiche che stabilizzano ulteriormente la molecola. Ciascuna catena ha un’estremità 3’ e un’estremità 5’.
Le due catene della doppia elica sono disposte in modo antiparallelo in quanto i legami fosfodiesterici che uniscono i pentosi contigui hanno polarità opposte. In altre parole, in corrispondenza di ciascuna delle due estremità della doppia elica, il terminale 5’ di una catena si confronta con il terminale 3’ dell’altra catena.
Le due eliche differiscono quindi per polarità, per sequenza e per composizione in basi, ma sono perfettamente complementari, cioè la sequenza di una catena è completamente determinata da quella della catena opposta.
le due eliche

Le due eliche differiscono quindi per polarità, per sequenza e per composizione in basi, ma sono perfettamente complementari, cioè la sequenza di una catena è completamente determinata da quella della catena opposta.
Le due eliche parentali si srotolano e ciascuna fa da stampo per la sintesi di un’elica-figlia secondo le regole dell’appaiamento delle basi (A=T; C=G). L’elevata capacità informativa del DNA si spiega con la sequenza dei nucleotidi. Quando la cellula va incontro a divisione, la doppia elica si apre grazie all’azione di enzimi specifici. Ciascuna elica funge da stampo per una nuova catena di DNA, secondo le regole dell’appaiamento delle basi.
Il DNA è chimicamente molto stabile, mentre l’RNA è molto suscettibile di degradazione. Durante ogni ciclo di divisione possono introdursi errori (tautomerie). Una timina tautomerica può legarsi ad un nucleotide che non sia l’adenina. La probabilità è di 1/1000. Il DNA umano è formato da 3 miliardi di nucleotidi.
una serie di articoli su nature
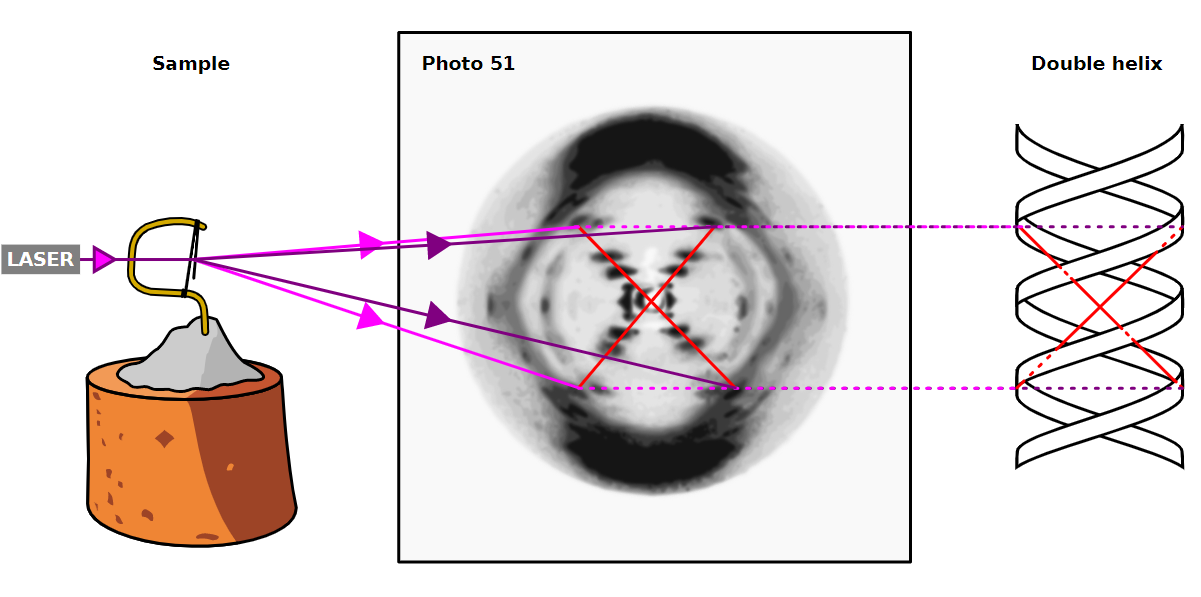
Il modello prevedeva inoltre che le due eliche fossero antiparallele e che la replicazione del DNA fosse di tipo semi-conservativa (un filamento vecchio e uno nuovo). Le evidenze sperimentali a supporto del loro modello furono riportate in una serie di cinque articoli pubblicati sullo stesso numero di “Nature” del 25 aprile 1953.
Tra questi articoli figurava anche quello di Rosalind Franklin e di Raymond Gosling (1926 – 2015), che conteneva i dati di diffrazione a raggi X, fondamentale per sostenere il modello e un articolo sulla struttura del DNA scritto da Maurice Wilkins. Per questa loro scoperta Watson e Crick ricevettero nel 1962 il Premio Nobel per la Medicina insieme a Wilkins.
F. Crick era, allora, un promettente fisico inglese che interruppe la sua carriera quando scoppiò la seconda guerra mondiale per unirsi all’Ammiragliato. Durante quegli otto anni, rivolse il suo campo di ricerca verso la biologia, dopo aver letto il libro dell’eminente fisico Erwin Schrodinger, “What is Life?” e alla fine si accasò al “Cavendish Laboratory” di Cambridge.
tra la foto 51 ed il nobel per la medicina

M. Wilkins era, invece, nato nel deserto della Nuova Zelanda, figlio di un medico-ricercatore irlandese, ma visse dall’età di sei anni nel Regno Unito. Dopo aver conseguito il dottorato in Fisica, venne coinvolto nel “Progetto Manhattan” per sviluppare la prima bomba nucleare negli Stati Uniti.
Anche in questo caso, il libro di Schrodinger lo ispirò a cercare la fisica nella genetica. Dopo la guerra, tornò in Inghilterra e si unì al laboratorio di biofisica di John Turton Randall (23marzo 1905 – 16giugno 1984) al King’s College di Londra. Questo divenne infine il luogo dove si creò il un team per individuare la struttura del DNA.
Il laboratorio includeva anche R. E. Franklin, l’eroina della biologia molecolare, una ricercatrice incredibilmente talentuosa che si era unita al Randall per aggiornare l’unità di cristallografia a raggi X.
Franklin e Wilkins

Essa analizzò insieme a M. Wilkins agli inizi degli anni ‘50 i cristalli di DNA con il metodo della diffrazione dei raggi X. I due videro che I polimeri di DNA erano elicoidali con due periodicità lungo il loro asse: una primaria di 3,4 Å e una secondaria di 34 Å.
La Franklin era figlia di un banchiere che aveva deciso all’età di quindici anni di diventare scienziata. All’età di diciotto anni era entrata a Cambridge causando una crisi familiare: il padre, convinto che il posto di una donna non fosse in laboratorio né all’università, aveva rifiutato di pagare l’iscrizione.
A 26 anni, Rosalind aveva pubblicato già cinque articoli e completato il dottorato. Cominciò a lavorare alla diffrazione a raggi X e rivelò subito abilità e ingegno da fuoriclasse. Fu fra i primi scienziati ad utilizzare i raggi X non per studiare cristalli ma materiali complessi come le molecole biologiche.
L’incontro di Watson con il gruppo del King’s College

James Watson era , invece, un brillante giovane scienziato americano che incontrò Wilkins per la prima volta ad una conferenza a Napoli nel 1951. L’anno precedente, Wilkins e uno dei suoi studenti di dottorato, Raymond Gosling, avevano prodotto diverse immagini a raggi X della struttura cristallina della molecola di DNA. Alcuni di questi furono presentati alla conferenza. Un’immagine in particolare si rivelò così intrigante per James Watson che si unì al Cavendish Laboratory lo stesso anno per perseguire la struttura del DNA. Qui incontrò F. Crick, e da questo momento le loro strade si unirono per lavorare insieme sul DNA.
Allo stesso tempo, anche R. Franklin si è unita alla corsa per interpretare la molecola elusiva. Nonostante lavorasse da sola, ella è arrivata sorprendentemente vicina a risolvere il puzzle. Un anno prima che F. Crick e J. Watson si occupassero del DNA , R. Franklin entrò al King’s College di Londra, sotto la supervisione di M. Wilkins.
wilkins e franklin: un rapporto difficile
|
|
|


I due, già da subito, non si piacquero affatto: Wilkins era timido e riflessivo mentre R. Franklin era diretta. Ella considerò sempre lo studio del DNA il proprio progetto di ricerca mentre Wilkins al contrario, ritenne che il DNA era affar suo e la trattò come una sua assistente. Dall’altra parte, ella considerò, invece, M. Wilkins un ricercatore “middle-class“ ed i suoi colleghi “repellenti”. Una volta , ella scrisse:
“mi sono organizzata in maniera da non vedere nessuno di loro quasi mai. Tra loro non c’è alcuna mente di prima categoria, e neanche una semplicemente buona. In pratica non trovo nessuno col quale poter desiderare di intavolare una discussione, scientifica o su altri argomenti”.
Discutere coi maschi, d’altronde in quel tempo, non era semplice. Le donne del laboratorio non erano ammesse a mangiare con i colleghi in sala mensa.
IL LAVORO DELLA FRANKLIN

Nel laboratorio la Franklin modificò i propri macchinari e riuscì a ottenere un fascio estremamente sottile di raggi X e fu capace, con i suoi studenti, di produrre campioni minuscoli di DNA e di sistemarli in fibre perfettamente paralleli.
Scoprì che la molecola assumeva forme diverse in funzione dell’umidità dell’aria. Mostrò che la molecola di fosfato che fa da impalcatura alla struttura doveva essere sul lato esterno del DNA e che, almeno nella forma “umida” , il DNA doveva avere forma elicoidale: una specie di scala a chiocciola. Tuttavia, rigorosa e prudente all’eccesso, decide di non pubblicare nulla prima di avere prove conclusive.
Nel 1951 Watson assistette a un seminario della Franklin ed ebbe l’opportunità di osservare le sue foto e di ascoltarla. Per sua stessa ammissione, ammise di aver capito poco. Tornò a Cambridge con due consapevolezze: 1) non gli piacque l’aspetto fisico né lo stile della Franklin e 2) che il DNA doveva avere qualcosa a che fare con un’elica.
INSEGUENDO IL MODELLO

Watson e Crick, allora, incominciarono a costruire in una settimana un modellino di DNA che risultò in tre filamenti, il fosforo sul lato interno e le basi azotate che spuntavano fuori dalla molecola. Tale modello venne poi spedito alla Franklin per un consiglio ma che lo bocciò come “inconsistente coi dati sperimentali”. In realtà era così sballato che il capo sezione dei due ricercatori intimò loro di lasciar perdere il DNA e trovare un argomento nel quale fossero competenti.
Ma pochi mesi dopo alcuni colpi di fortuna (la conoscenza di M. Wilkins che consegnò loro la famosa foto 51 fatta dalla Franklin, e a sua insaputa dove veniva mostrata la forma di un’elica e la dimensione esatta del diametro della molecola del DNA), di astuzia e di genio (le scoperte di E. Chargaff) destinarono a Watson e a Crick l’opportunità di ricevere il premio Nobel. Lo stesso Watson scriverà in seguito:
“Nell’istante in cui vidi quell’immagine rimasi a bocca aperta. Molti dei parametri vitali dell’elica erano lì “.
Anche Crick ammetterà, più tardi:
“senza quei dati la formulazione del nostro modello sarebbe stata altamente improbabile, se non impossibile “.
IL LAVORO DELLA FRANKLIN SFRUTTATO DA WATSON E CRICK
La fotografia della Franklin, disegnata da Watson, fu utilizzata da Crick per calcolare alcuni parametri dell’elica. Si trattava di una fotografia di diffrazione a raggi X della forma B di fibre di DNA purificato.
Gli archi marcati sulla periferia mostravano aspetti della molecola strettamente spaziati che fornivano una stima della periodicità delle basi azotate distanziate di 3,4 A (angstrom). Il profilo interno di macchie a croce mostrava l’aspetto più grossolano della molecola indicante la sua natura a elica. Il DNA era precipitato con una soluzione concentrata di etanolo e formava delle fibre in cui le eliche si impacchettavano a formare delle strutture simili a cristalli (cristalliti) che potevano essere analizzate mediante diffrazione con raggi X. Nel 1962, dopo la morte della Franklin (a causa di un tumore provocato, probabilmente, dalle alte dosi di raggi X a cui si era esposta nel corso dei suoi esperimenti), Watson, Crick e Wilkins ricevettero congiuntamente il Premio Nobel per la medicina e la doppia elica contribuì a portare la molecola del DNA alla ribalta.
un nobel contestato

Dal momento che la scoperta del modello si basò essenzialmente sui dati della Franklin, ancora oggi esistono pareri molto eterogenei nella comunità scientifica su chi avrebbe dovuto ricevere tale premio.
Watson e Crick elaborarono vari aspetti teorici della struttura del DNA. Tuttavia, per confermare la sua accuratezza, le chiare immagini a raggi X della Franklin e le sue misurazioni accurate di varie distanze all’interno della cellula unitaria del DNA si rivelarono cruciali. Incorporando questi dati, essi riuscirono a pubblicare la struttura a doppia elica del DNA nel 1953, oltre a mostrare la sua capacità di replicazione. Nonostante il ruolo chiave svolto dal lavoro della Franklin, quest’ultima venne omessa dalla citazione del Premio Nobel.
L’ERRORE DI PAULING
Tuttavia, l’idea che la molecola del DNA fosse costituita da un’elica non era affatto nuova: già il chimico statunitense Pauling, vincitore di ben due premi Nobel (per la chimica nel 1954 e per la pace nel 1962 ), aveva annunciato proprio nel 1953 un modello a tripla elica con i gruppi fosfato all’interno.
Incredibilmente, egli scrisse un libro sulla natura del legame chimico non considerando che le forze repulsive generate dalle cariche negative dei gruppi fosfati avrebbero fatto collassare la struttura.
Anche lo stesso Wilkins era convinto che si trattasse di un’elica, e cercò di determinarla non mediante modelli come Watson e Crick ma attraverso la diffrazione a raggi X. Le fotografie del suo laboratorio fornirono una conferma della struttura e Wilkins condivise con loro il premio Nobel nel 1962.
clicca per andare alla seconda parte
Bibliografia:
- Dahm R. (2005). “Friedrich Miescher and the discovery of DNA.” Developmental Biology 278: 274-288
- Avery O.T., MacLeod C.M., McCarty M.(1944) “Studies of the chemical nature of the substance inducing transformation of pneumococcal types. Induction of transformation by a deoxyribonucleic acid fraction isolated from pneumococcus type III “ J. Exp. Med.,79: 137-158
- Hershey A.D., Chase M. (1952) “Independent functions of viral proteins and nucleic acid in growth of bacteriophage” J. Gen. Physiol. 36 : 39-56
- Watson J.D. & Crick F.H.C. (1953) “A Structure for Deoxyribose Nucleic Acid“. Nature 171, 737–738